
Geschrieben vom Qubic Scientific Team: Verzweigungs-Verhältnis und Kritikalität in biologischen Netzwerken, in künstlichen Netzwerken und als bioinspiriertes Prinzip in Neuraxon.
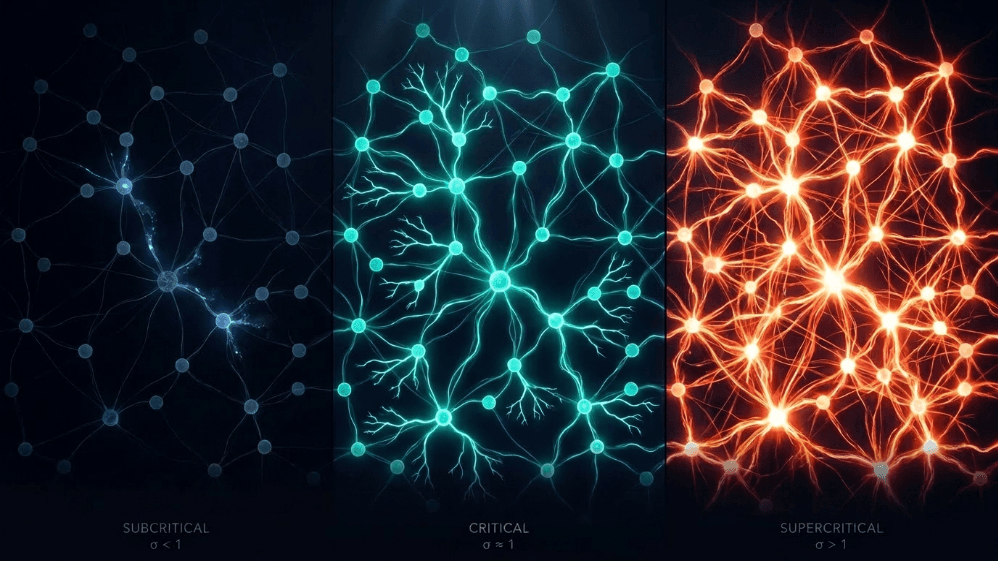
Was haben eine Schneelawine, ein Waldbrand, ein Erdbeben und die spontane Aktivität des Großhirnrinde gemeinsam?
Sie teilen alle eine Grenze zwischen Ordnung und Chaos, was als kritischer Zustand bezeichnet wird. Im Gehirn wird diese Grenze durch einen einfachen Parameter gemessen: das Verzweigungs-Verhältnis (σ oder m). Es wäre etwas wie das durchschnittliche Verhältnis der neuronalen "Nachkommen", die jede "Eltern"-Neuron aktiviert. Wenn σ ≈ 1, stirbt die Aktivität weder aus noch explodiert sie; sie hallt wider.
Beggs und Plenz (2003) zeichneten die spontane Aktivität des Gehirns bei Ratten auf und stellten fest, dass die Aktivität kaskadenartige Muster bildete, die sogenannten neuronalen Lawinen, mit einem Verzweigungs-Verhältnis nahe 1. Das Gehirn schien an einem kritischen Punkt zu leben. Bei Menschen erscheint das Verzweigungs-Verhältnis σ erneut nahe der Eins (Wang et al., 2025; Plenz et al., 2021; Wilting & Priesemann, 2019).
Am kritischen Punkt zeigen Systeme gleichzeitig maximale Sensitivität gegenüber Störungen (Reaktionsfähigkeit), maximale dynamische Kapazität (Anzahl der zugänglichen Zustände), maximale Informationsübertragung und maximale Komplexität (Timme et al., 2016; Shew et al., 2009, 2011).
Was ist das Verzweigungs-Verhältnis und wie wird es gemessen?
Konzeptionell ist das Verzweigungs-Verhältnis trivial: Wenn es zu einem Zeitpunkt t A(t) aktive Neuronen gibt und zu t+1 A(t+1), dann:
σ = ⟨ A(t+1) / A(t) ⟩
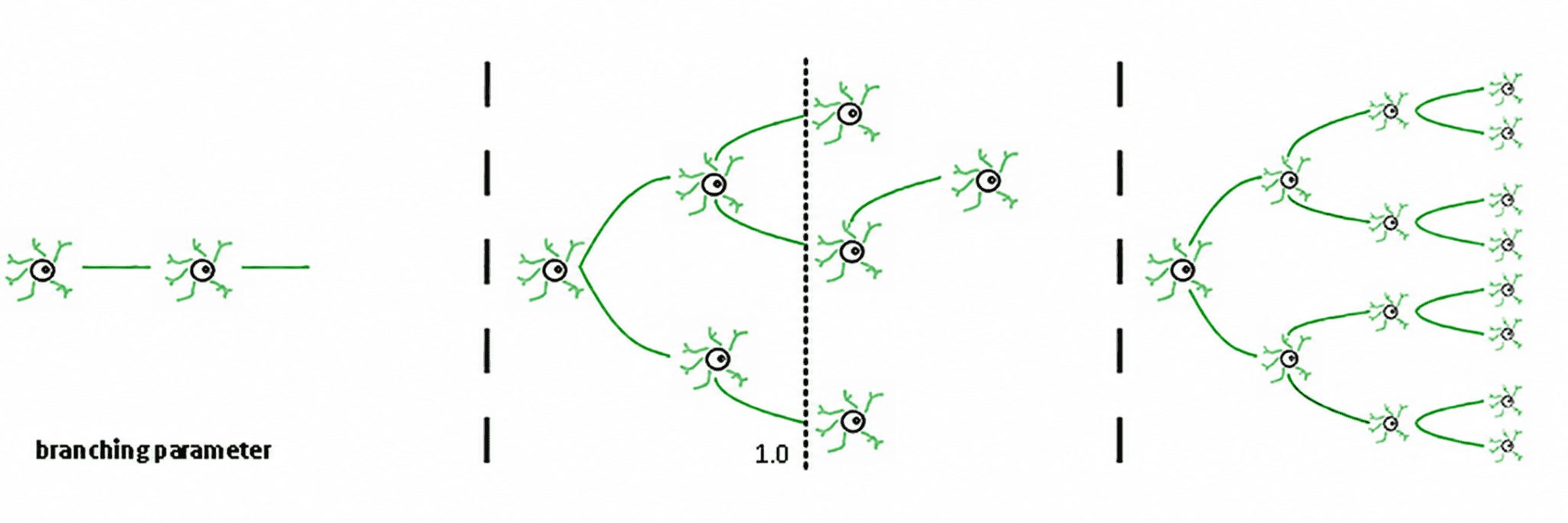
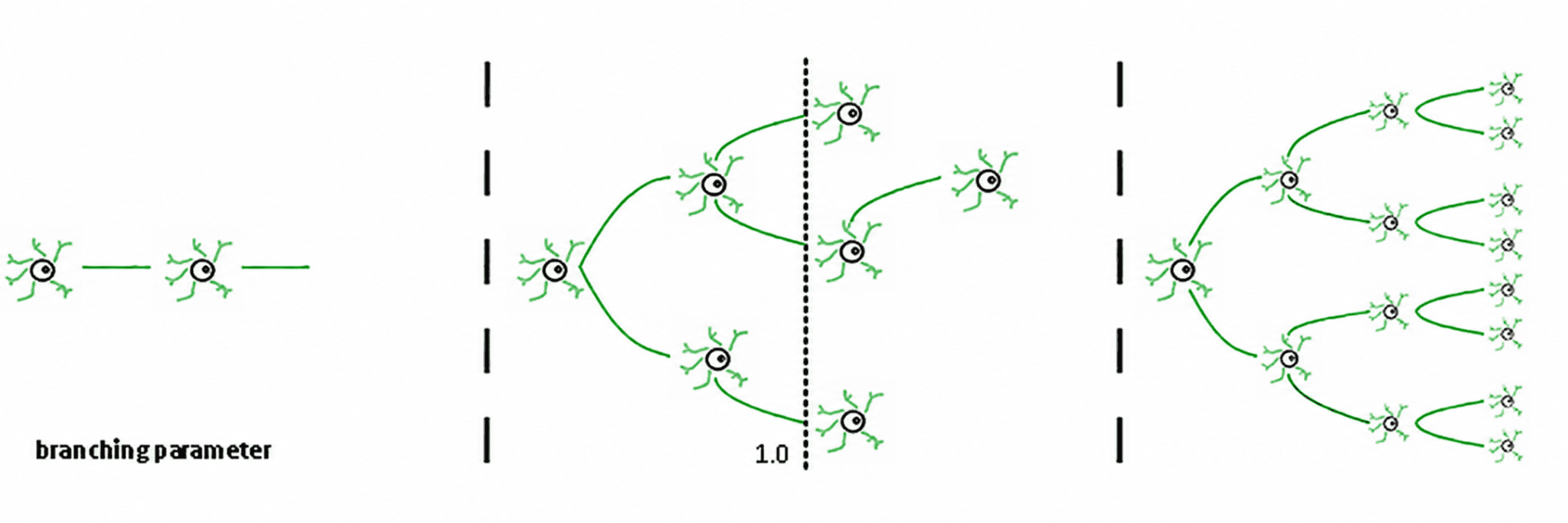
Drei Regime ergeben sich daraus (de Carvalho & Prado, 2000; Haldeman & Beggs, 2005):
Subkritisch (σ < 1): Aktivität nimmt ab; das System "vergisst" die Störung schnell. Es ist stabil, aber arm an Gedächtnis und nicht sehr ausdrucksstark.
Superkritisch (σ > 1): Aktivität explodiert in Kaskaden. Dies ist das Zeichen pathologischer Regime wie epileptischer Anfälle (Hsu et al., 2008; Hagemann et al., 2021).
Kritisch (σ ≈ 1): Jeder Spike erzeugt im Durchschnitt einen weiteren Spike. Aktivität widerhallt, neuronale Lawinen gehorchen Potenzgesetzen, und das System behält ein strukturiertes Gedächtnis der Eingabe.
Die Schönheit von σ ist, dass es eine einzelne Zahl ist, die das globale dynamische Regime zusammenfasst. Aber es zu messen, ist weniger trivial. Wenn es auf in vivo kortikale Aufzeichnungen angewendet wird, zeigt die Messung, dass der Cortex nicht genau bei σ = 1 operiert, sondern leicht darunter, in einem Regime, das die Autoren als widerhallend bezeichnen (Wilting et al., 2018). Der Unterschied ist wichtig: Genau bei σ = 1 zu sein, wäre wie das Fahren eines Fahrrads, das auf einem Drahtseil balanciert; leicht darunter erlaubt eine schnelle Anpassung an die Anforderungen der Aufgabe, ohne das Risiko einer unkontrollierbaren Explosion.
Kritikalität in künstlichen neuronalen Netzwerken: Vom Rand des Chaos bis zur Reservoir Computing
Bertschinger und Natschläger (2004) zeigten, dass zufällige rekurrente Schwellenwertnetzwerke ihre maximale Rechenkapazität bei zeitlichen Verarbeitungsaufgaben genau an der Übergangsfläche zwischen Ordnung und Chaos erreichen.
Boedecker et al. (2012) erweiterten die Analyse auf Echo-State-Netzwerke im Reservoir-Computing-Paradigma und bestätigten, dass die Informationsübertragungskapazität und das aktive Gedächtnis am Rand des Chaos maximiert sind.
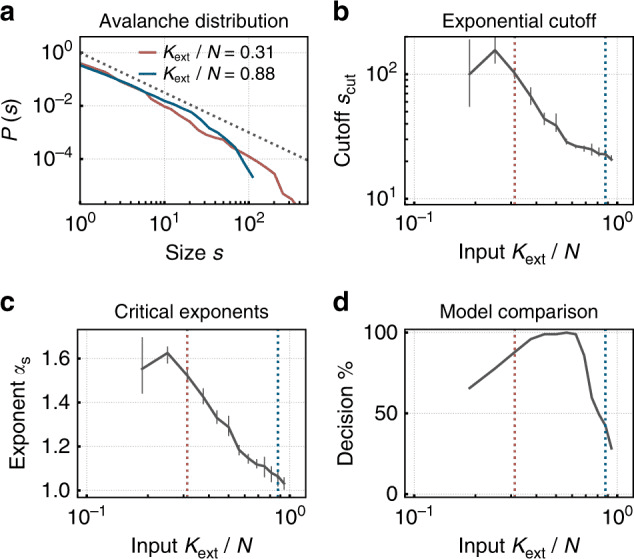
Abb. 3. Ein spiking neuromorphes Netzwerk mit synaptischer Plastizität selbstorganisiert sich in Richtung Kritikalität unter niedrigem externen Input und weist eine Größe der Lawinenverteilung auf, die Potenzgesetze gehorcht — das Markenzeichen des kritischen Zustands in biologischen und künstlichen neuronalen Netzwerken. Bei höherem Input wechselt das Netzwerk in ein subkritisches Regime mit abgeschnittenen Verteilungen. Reproduziert aus Cramer et al. (2020), Nature Communications, 11, 2853. CC BY 4.0.
In der Sprache der künstlichen neuronalen Netzwerke wird der Messparameter als spektraler Radius bezeichnet. Wenn er 1 überschreitet, divergieren die Trajektorien exponentiell (Chaos); wenn er deutlich unter 1 liegt, kollabiert das Netzwerk zum Fixpunkt und verliert Gedächtnis. Der spektrale Radius, der nahe 1 liegt, ist in diesem Kontext das formale Äquivalent zu biologischen σ ≈ 1 (Magnasco, 2022; Morales et al., 2023). In spiking neuronalen Netzwerken kann das Verzweigungs-Verhältnis mit Methoden gemessen werden, die fast identisch sind mit denen, die in neuronalen Kulturen verwendet werden (Cramer et al., 2020; Zeraati et al., 2024).
Warum maximiert die Gehirnkritikalität die neuronale Berechnung?
In der Nähe von σ ≈ 1 zu arbeiten, bietet vier Vorteile, die sowohl für die Hypothese des kritischen Gehirns als auch für das Design gehirn-inspirierter KI-Systeme zentral sind:
Maximaler dynamischer Bereich. Shew et al. (2009) zeigten, dass der Bereich der Eingangsintensitäten, den der Cortex diskriminieren kann, maximal ist, wenn das Gleichgewicht zwischen Erregung und Hemmung das Netzwerk in den Zustand der Kritikalität versetzt.
Maximierte Informationskapazität. Die Entropie von Lawinenmustern und die wechselseitige Information zwischen Eingabe und Ausgabe erreichen bei σ ≈ 1 ihren Höhepunkt (Shew et al., 2011).
Optimale Vergänglichkeit des Gedächtnisses. Im kritischen Regime wird die Störung nur lange genug aufrechterhalten, um die Verarbeitung zu beeinflussen, ohne die ferne Zukunft zu kontaminieren; es ist der süße Punkt zwischen Stabilität und zeitlicher Integration (Boedecker et al., 2012).
Komplexität als einheitliches Maß. Timme et al. (2016) zeigten, dass die neuronale Komplexität genau am kritischen Punkt maximiert ist und Kritikalität mit formalen Theorien des Bewusstseins und der Verarbeitung verknüpft.
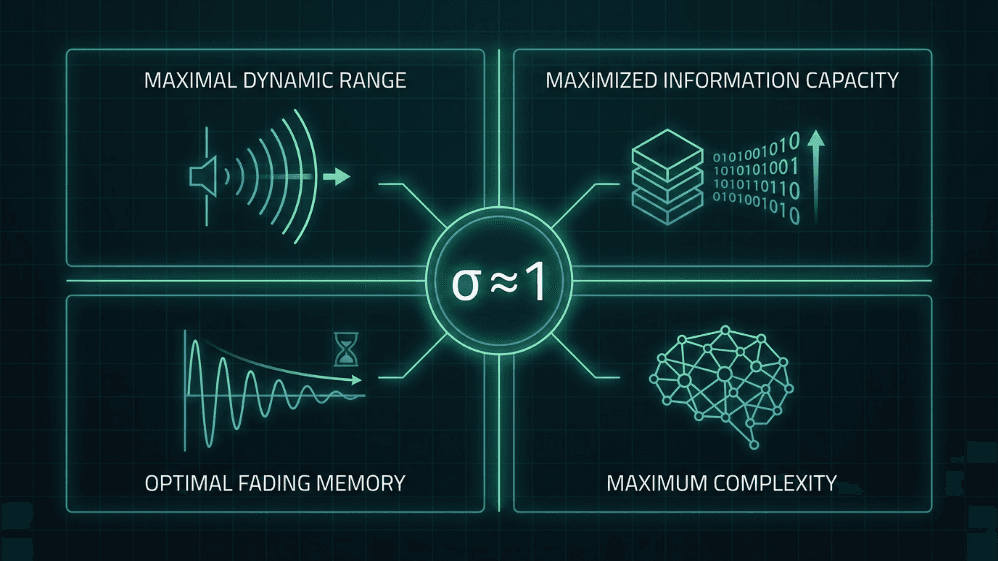
Abb. 4. Vier berechnungstechnische Vorteile des Betriebs in der Nähe des kritischen Verzweigungs-Verhältnisses (σ ≈ 1). Bei Kritikalität erreichen neuronale Netzwerke maximalen dynamischen Bereich, maximierte Informationskapazität, optimales Vergessen des Gedächtnisses und maximale Komplexität — Eigenschaften, die sowohl für die Hypothese des kritischen Gehirns als auch für das Design gehirn-inspirierter KI zentral sind.
Das Gehirn operiert nicht immer bei σ = 1
Das bedeutet nicht, dass das Gehirn immer bei σ = 1 operiert. Beweise deuten vielmehr auf ein leicht subkritisches und modulierbares Regime hin: Bei anspruchsvollen Aufgaben nähert sich das Netzwerk der Kritikalität, während es sich im Tiefschlaf davon entfernt, und pathologische Zustände (Epilepsie, tiefe Anästhesie, bestimmte psychiatrische Zustände) sind mit messbaren Abweichungen von diesem Betriebsbereich verbunden (Meisel et al., 2017; Zimmern, 2020). Das Verzweigungs-Verhältnis wird zu einem dynamischen Biomarker des funktionalen Zustands des Nervensystems.
Warum wir das Verzweigungs-Verhältnis in Neuraxon verwenden: Bioinspirierte KI-Entwicklung am Rand des Chaos
Neuraxon ist ein bioinspiriertes System, das dynamische Prinzipien des Cortex als Designbeschränkungen übernimmt. Das Verzweigungs-Verhältnis ist eines der wichtigsten, und wir verwenden es aus vier Gründen:
Als Echtzeit-Betriebsinvarianz für die Stabilität neuronaler Netzwerke
In tiefen spiking oder rekurrenten Architekturen ist das doppelte Risiko des Aktivitätskollapses (stilles Netzwerk, verschwindende Gradienten) und der unkontrollierbaren Explosion (Sättigung, explodierende Gradienten) strukturell. Die Überwachung von σ in Echtzeit gibt uns einen einzigen diagnostischen Skalar, unabhängig von der konkreten Architektur, der anzeigt, ob das System im rechnerischen Sinne lebendig ist.
Als bioinspiriertes Selbstregulationsziel durch selbstorganisierte Kritikalität
Das Netzwerk organisiert sich selbst in Richtung Kritikalität, ohne dass eine zentrale Feinabstimmung erforderlich ist, und repliziert das Prinzip der selbstorganisierten Kritikalität (Bornholdt & Röhl, 2003; Levina et al., 2007). Dies reduziert die Sensitivität gegenüber Hyperparametern drastisch und verleiht dem System Robustheit gegenüber Verteilungverschiebungen. Wie wir in NIA Band 7 über künstliches Leben und digitale Ökosysteme erkundet haben, ist dies genau, wie emergente Komplexität aus lokalen Regeln ohne zentrale Kontrolle entsteht.
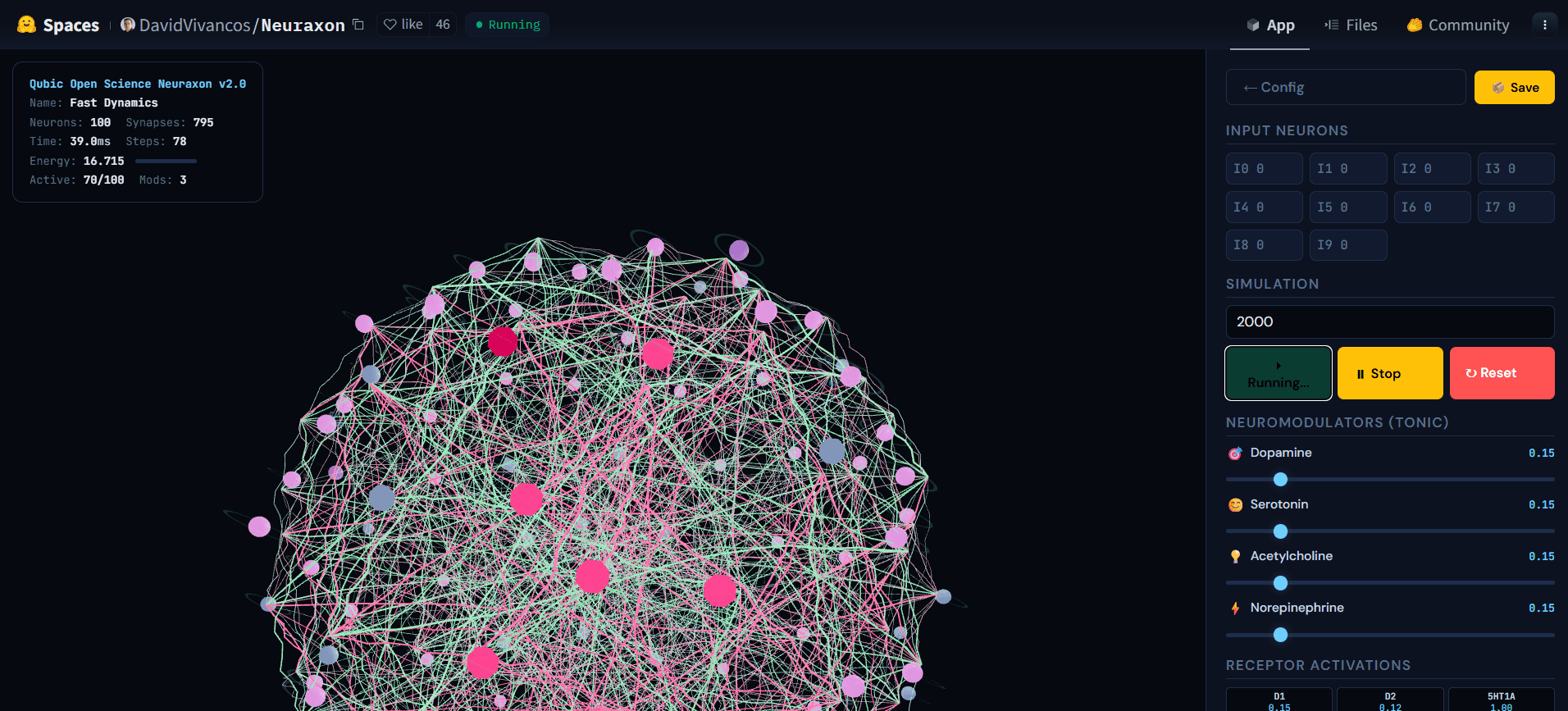
Abb. 5. Neuraxon 3D-Netzwerk während aktiver Simulation, das kaskadierende Aktivität über ternäre Zustandsneuronen zeigt. Helle aktive Knoten (pink) propagieren Signale durch exzitatorische (grün) und inhibitorische (pink) Verbindungen, während andere Neuronen inaktiv bleiben (grau) und ein widerhallendes Regime in der Nähe des kritischen Verzweigungs-Verhältnisses (σ ≈ 1) veranschaulichen. Dieser ausgewogene Zustand — weder still noch explosiv — ist das, worauf Neuraxon sich selbst organisiert, indem es bioinspirierte Kritikalitätsprinzipien verwendet. Erforschen Sie die interaktive Demo unter huggingface.co/spaces/DavidVivancos/Neuraxon. Quelle: Qubic Scientific Team.
Als Brücke zwischen neurowissenschaftlicher Beobachtung und KI-Design
Das Verzweigungs-Verhältnis ist eine der sehr wenigen Größen, die mit dem gleichen Formalismus in der Elektrophysiologie, fMRI und künstlichen Netzwerken gemessen werden. Dies ermöglicht das Testen bidirektionaler Hypothesen: Wenn eine Intervention die biologische Kritikalität verbessert, können wir fragen, ob dieselbe Intervention — in die künstliche Architektur übersetzt — die Berechnung des Modells verbessert und umgekehrt. Dieses Prinzip ist zentral für den Neuromodulationsrahmen und die astrozytischen Gating-Mechanismen, die wir in früheren Bänden dieser Akademie entwickelt haben.
Als funktionales, nicht ästhetisches Kriterium für gehirn-inspirierte KI
Kritikalität ist eine operationale Einschränkung mit empirischen Konsequenzen. Arbeiten in der Nähe des widerhallenden Regimes verbessert — wie in unseren internen Bewertungen und eingereichten Publikationen gemessen — die Generalisierungsfähigkeit, die Stabilität unter Eingabestörungen, die darstellende Fülle und die temporale Kohärenz des Denkens. Diese Effekte stimmen qualitativ mit den in der biologischen (Cocchi et al., 2017) und künstlichen (Cramer et al., 2020; Morales et al., 2023) Literatur berichteten überein.
Das Verzweigungs-Verhältnis: Von der statistischen Physik zur gehirn-inspirierten KI-Architektur
Das Verzweigungs-Verhältnis ist eines dieser konzeptionellen rara avis: einfach genug, um auf eine einzige Formel reduziert zu werden, tief genug, um statistische Physik, Neurowissenschaften, KI und Systemdesign zu überbrücken. Für das biologische Gehirn scheint σ ≈ 1 das Regime zu sein, in dem die tugendhafte Kombination aus Sensitivität, Gedächtnis, Ausdruckskraft und Robustheit entsteht. Für künstliche Netzwerke sagt dasselbe Grenzgebiet — umbenannt als der Rand des Chaos — die maximale Berechnungskapazität vorher.
Und für Neuraxon ist es ein Leitprinzip des bioinspirierten Designs: eine überprüfbare, selbstregulierende und biologisch bedeutende Metrik, die uns hilft, das System am Leben zu halten, im reichhaltigsten Sinne des Wortes.
Literaturverzeichnis
Beggs, J. M., & Plenz, D. (2003). Neuronale Lawinen in neokortikalen Schaltkreisen. Das Journal of Neuroscience, 23(35), 11167–11177. https://doi.org/10.1523/JNEUROSCI.23-35-11167.2003
Bertschinger, N., & Natschläger, T. (2004). Echtzeitberechnung am Rand des Chaos in rekurrenten neuronalen Netzwerken. Neural Computation, 16(7), 1413–1436. https://doi.org/10.1162/089976604323057443
Boedecker, J., Obst, O., Lizier, J. T., Mayer, N. M., & Asada, M. (2012). Informationsverarbeitung in Echo-State-Netzwerken am Rand des Chaos. Theorie in Biosciences, 131(3), 205–213. https://doi.org/10.1007/s12064-011-0146-8
Bornholdt, S., & Röhl, T. (2003). Selbstorganisierte kritische neuronale Netzwerke. Physikalische Überprüfung E, 67(6), 066118. https://doi.org/10.1103/PhysRevE.67.066118
Cocchi, L., Gollo, L. L., Zalesky, A., & Breakspear, M. (2017). Kritikalität im Gehirn: Eine Synthese von Neurobiologie, Modellen und Kognition. Fortschritte in der Neurobiologie, 158, 132–152. https://doi.org/10.1016/j.pneurobio.2017.07.002
Cramer, B., Stöckel, D., Kreft, M., Wibral, M., Schemmel, J., Meier, K., & Priesemann, V. (2020). Kontrolle von Kritikalität und Berechnung in spiking neuromorphen Netzwerken mit Plastizität. Nature Communications, 11, 2853. https://doi.org/10.1038/s41467-020-16548-3
de Carvalho, J. X., & Prado, C. P. C. (2000). Selbstorganisierte Kritikalität im Olami-Feder-Christensen-Modell. Physikalische Überprüfung Briefe, 84(17), 4006–4009. https://doi.org/10.1103/PhysRevLett.84.4006
Derrida, B., & Pomeau, Y. (1986). Zufällige Netzwerke von Automaten: Eine einfache geglättete Annäherung. Europhysics Letters, 1(2), 45–49. https://doi.org/10.1209/0295-5075/1/2/001
Hagemann, A., Wilting, J., Samimizad, B., Mormann, F., & Priesemann, V. (2021). Bewertung der Kritikalität in der Einzelneuronenaktivität des menschlichen epileptischen Cortex vor Anfällen. PLOS Computational Biology, 17(3), e1008773. https://doi.org/10.1371/journal.pcbi.1008773
Haldeman, C., & Beggs, J. M. (2005). Kritisches Verzweigen erfasst die Aktivität in lebenden neuronalen Netzwerken und maximiert die Anzahl metastabiler Zustände. Physikalische Überprüfung Briefe, 94(5), 058101. https://doi.org/10.1103/PhysRevLett.94.058101
Hsu, D., Chen, W., Hsu, M., & Beggs, J. M. (2008). Eine offene Hypothese: Ist Epilepsie erlernt, und kann sie verlernt werden? Epilepsie & Verhalten, 13(3), 511–522. https://doi.org/10.1016/j.yebeh.2008.05.007
Langton, C. G. (1990). Berechnung am Rand des Chaos: Phasenübergänge und emergente Berechnung. Physica D: Nichtlineare Phänomene, 42(1–3), 12–37. https://doi.org/10.1016/0167-2789(90)90064-V
Levina, A., Herrmann, J. M., & Geisel, T. (2007). Dynamische Synapsen, die selbstorganisierte Kritikalität in neuronalen Netzwerken verursachen. Nature Physics, 3(12), 857–860. https://doi.org/10.1038/nphys758
Magnasco, M. O. (2022). Robustheit und Flexibilität neuronaler Funktionen durch dynamische Kritikalität. Entropie, 24(5), 591. https://doi.org/10.3390/e24050591
Meisel, C., Klaus, A., Vyazovskiy, V. V., & Plenz, D. (2017). Das Zusammenspiel zwischen kurz- und langfristigen zeitlichen Korrelationen prägt die Dynamik des Cortex über Wachheitszustände hinweg. Das Journal of Neuroscience, 37(42), 10114–10124. https://doi.org/10.1523/JNEUROSCI.0448-17.2017
Morales, G. B., di Santo, S., & Muñoz, M. A. (2023). Enthüllung der intrinsischen Dynamik biologischer und künstlicher neuronaler Netzwerke: Von Kritikalität zu optimalen Darstellungen. Frontiers in Complex Systems, 1, 1276338. https://doi.org/10.3389/fcpxs.2023.1276338
Plenz, D., Ribeiro, T. L., Miller, S. R., Kells, P. A., Vakili, A., & Capek, E. L. (2021). Selbstorganisierte Kritikalität im Gehirn. Frontiers in Physics, 9, 639389. https://doi.org/10.3389/fphy.2021.639389
Shew, W. L., Yang, H., Petermann, T., Roy, R., & Plenz, D. (2009). Neuronale Lawinen implizieren maximalen dynamischen Bereich in kortikalen Netzwerken bei Kritikalität. Das Journal of Neuroscience, 29(49), 15595–15600. https://doi.org/10.1523/JNEUROSCI.3864-09.2009
Shew, W. L., Yang, H., Yu, S., Roy, R., & Plenz, D. (2011). Die Informationskapazität und -übertragung sind in ausgewogenen kortikalen Netzwerken mit neuronalen Lawinen maximiert. Das Journal of Neuroscience, 31(1), 55–63. https://doi.org/10.1523/JNEUROSCI.4637-10.2011
Spitzner, F. P., Dehning, J., Wilting, J., Hagemann, A., Neto, J. P., Zierenberg, J., & Priesemann, V. (2021). MR. Estimator, ein Werkzeug zur Bestimmung intrinsischer Zeitskalen aus subabgetasteten Spike-Aktivitäten. PLOS ONE, 16(4), e0249447. https://doi.org/10.1371/journal.pone.0249447
Timme, N. M., Marshall, N. J., Bennett, N., Ripp, M., Lautzenhiser, E., & Beggs, J. M. (2016). Kritikalität maximiert die Komplexität im neuronalen Gewebe. Frontiers in Physiology, 7, 425. https://doi.org/10.3389/fphys.2016.00425
Turrigiano, G. G. (2008). Der selbstabstimmende Neuron: Synaptische Skalierung von exzitatorischen Synapsen. Cell, 135(3), 422–435. https://doi.org/10.1016/j.cell.2008.10.008
Wang, J., Cao, R., Brunton, B. W., Smith, R. E. W., Buckner, R. L., & Liu, T. T. (2025). Genetische Beiträge zur kritikalität des Gehirns und deren Beziehung zu menschlichen kognitiven Funktionen. Proceedings of the National Academy of Sciences, 122(26), e2417010122. https://doi.org/10.1073/pnas.2417010122
Wilting, J., Dehning, J., Pinheiro Neto, J., Rudelt, L., Wibral, M., Zierenberg, J., & Priesemann, V. (2018). Das Arbeiten in einem widerhallenden Regime ermöglicht eine schnelle Anpassung der Netzwerkzustände an die Anforderungen der Aufgabe. Frontiers in Systems Neuroscience, 12, 55. https://doi.org/10.3389/fnsys.2018.00055
Wilting, J., & Priesemann, V. (2018). Kollektive dynamische Zustände aus weitgehend unbeobachteten Systemen ableiten. Nature Communications, 9, 2325. https://doi.org/10.1038/s41467-018-04725-4
Wilting, J., & Priesemann, V. (2019). 25 Jahre Kritikalität in der Neurowissenschaft — Etablierte Ergebnisse, offene Kontroversen, neue Konzepte. Aktuelle Meinung in Neurobiologie, 58, 105–111. https://doi.org/10.1016/j.conb.2019.08.002
Yu, C. (2022). Auf dem Weg zu einer einheitlichen Analyse der Hypothese der Gehirnkritikalität: Überprüfung mehrerer verfügbarer Werkzeuge. Frontiers in Neural Circuits, 16, 911245. https://doi.org/10.3389/fncir.2022.911245
Zeraati, R., Engel, T. A., & Levina, A. (2024). Schätzung von intrinsischen Zeitskalen und Kritikalität aus neuronalen Aufzeichnungen: Methoden und Fallstricke. Aktuelle Meinung in Neurobiologie, 86, 102871. https://doi.org/10.1016/j.conb.2024.102871
Zimmern, V. (2020). Warum die Gehirnkritikalität klinisch relevant ist: Eine Scoping-Studie. Frontiers in Neural Circuits, 14, 54. https://doi.org/10.3389/fncir.2020.00054
Erforschen Sie die vollständige Neuraxon Intelligence Academy
Dies ist Band 8 der #Neuraxon Intelligenz#academy des #Qubic Wissenschaftlichen Teams. Wenn Sie gerade erst dazustoßen, erkunden Sie die gesamte Serie, um ein vollständiges Verständnis der Wissenschaft hinter Neuraxon, #aigarth , und Qubics Ansatz zur gehirn-inspirierten, #decentralized künstlichen Intelligenz zu entwickeln:
NIA Vol. 1: Warum Intelligenz nicht in Schritten, sondern in Zeit berechnet wird — Erforscht, warum biologische Intelligenz in kontinuierlicher Zeit und nicht in diskreten Berechnungsschritten wie traditionelle LLMs arbeitet.
NIA Vol. 2: Ternäre Dynamik als Modell lebender Intelligenz — Erklärt ternäre Dynamik und warum Dreizustandslogik (exzitatorisch, neutral, inhibitorisch) für die Modellierung lebender Systeme wichtig ist.
NIA Vol. 3: Neuromodulation und gehirn-inspirierte KI — Behandelt Neuromodulation und wie die chemische Signalübertragung im Gehirn (Dopamin, Serotonin, Acetylcholin, Noradrenalin) die Architektur von Neuraxon inspiriert.
NIA Vol. 4: Neuronale Netzwerke in KI und Neurowissenschaften — Ein tiefer Vergleich zwischen biologischen neuronalen Netzwerken, künstlichen neuronalen Netzwerken und Neuraxons Ansatz der dritten Option.
NIA Vol. 5: Astrozyten und gehirn-inspirierte KI — Wie astrozytische Gating die Plastizität neuronaler Netzwerke durch den AGMP-Rahmen in Neuraxon transformiert.
NIA Vol. 6: Bewusste Maschinen vs. Intelligente Organismen: KI-Bewusstsein erklärt — Erforscht das KI-Bewusstsein durch die Linse der Global Workspace Theory, der integrierten Informationstheorie und der prädiktiven Kodierung.
NIA Vol. 7: Conways Spiel des Lebens, künstliches Leben und digitale Ökosysteme — Die Wissenschaft hinter Qubic, Aigarth und Neuraxons Ansatz zur entstehenden Komplexität und selbstorganisierten Kritikalität in dezentraler KI.
Qubic ist ein dezentrales, Open-Source-Netzwerk für experimentelle Technologie. Um mehr zu erfahren, besuchen Sie qubic.org. Beteiligen Sie sich an der Diskussion auf X, Discord und Telegram.