
Napisane przez Zespół Qubic Scientific Wskaźnik rozgałęzienia i krytyczność w sieciach biologicznych, w sieciach sztucznych oraz jako zasada bioinspirowana w Neuraxon
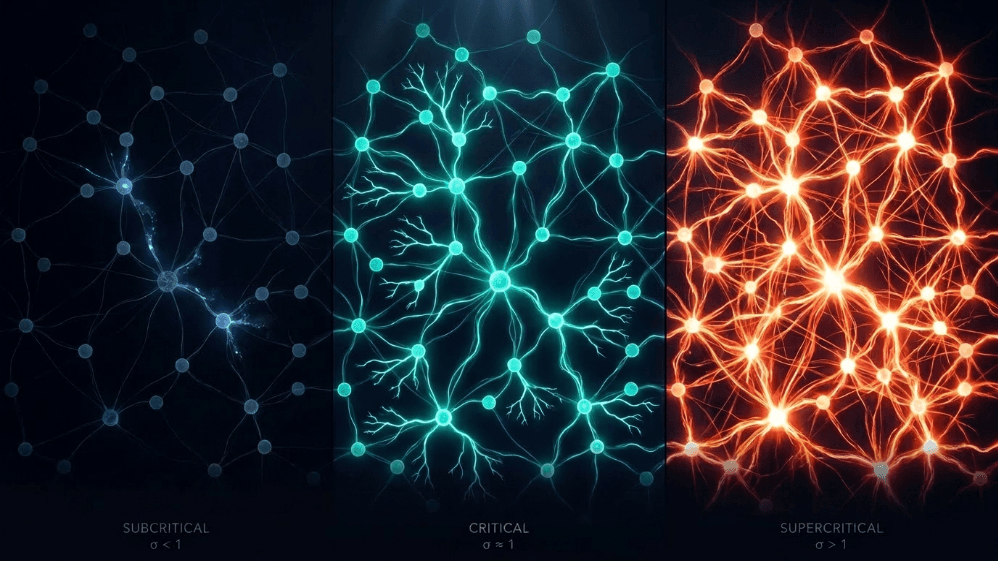
Co mają wspólnego lawina śnieżna, pożar lasu, trzęsienie ziemi i spontaniczna aktywność kory mózgowej?
Wszyscy dzielą granicę między porządkiem a chaosem, co nazywa się stanem krytycznym. W mózgu ta krawędź mierzona jest prostym parametrem: wskaźnikiem rozgałęzienia (σ lub m). To coś w rodzaju średniego wskaźnika "potomstwa" neuronów, które każdy neuron "rodzic" aktywuje. Gdy σ ≈ 1, aktywność ani nie zanika, ani nie eksploduje; odbija się.
Beggs i Plenz (2003) zarejestrowali spontaniczną aktywność kory mózgowej u szczurów i odkryli, że aktywność formowała wzory kaskadowe, tzw. lawiny neuronalne, z wskaźnikiem rozgałęzienia bliskim 1. Mózg wydawał się żyć na punkcie krytycznym. U ludzi wskaźnik rozgałęzienia σ ponownie wydaje się bliski jedności (Wang i in., 2025; Plenz i in., 2021; Wilting & Priesemann, 2019).
W punkcie krytycznym systemy jednocześnie wykazują maksymalną wrażliwość na perturbacje (reaktywność), maksymalną pojemność dynamiczną (liczba dostępnych stanów), maksymalną transmisję informacji oraz maksymalną złożoność (Timme i in., 2016; Shew i in., 2009, 2011).
Czym jest wskaźnik rozgałęzienia i jak jest mierzony?
Koncepcyjnie, wskaźnik rozgałęzienia jest trywialny: jeśli w momencie t jest A(t) aktywnych neuronów, a w t+1 jest A(t+1), to:
σ = ⟨ A(t+1) / A(t) ⟩
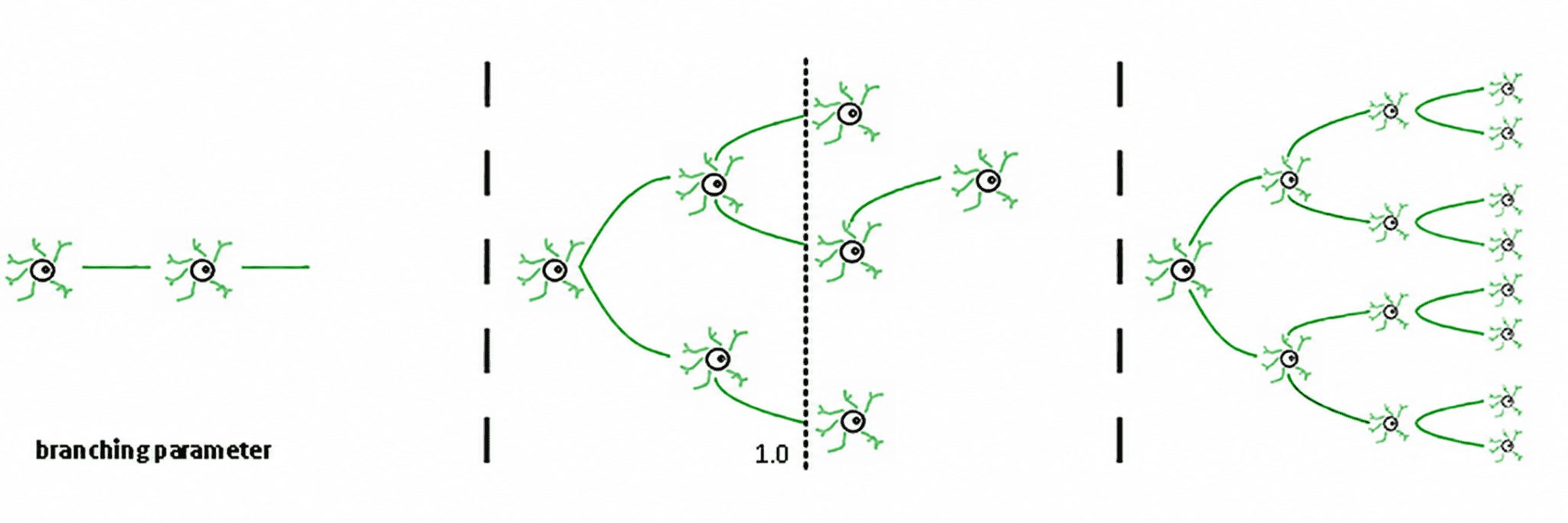
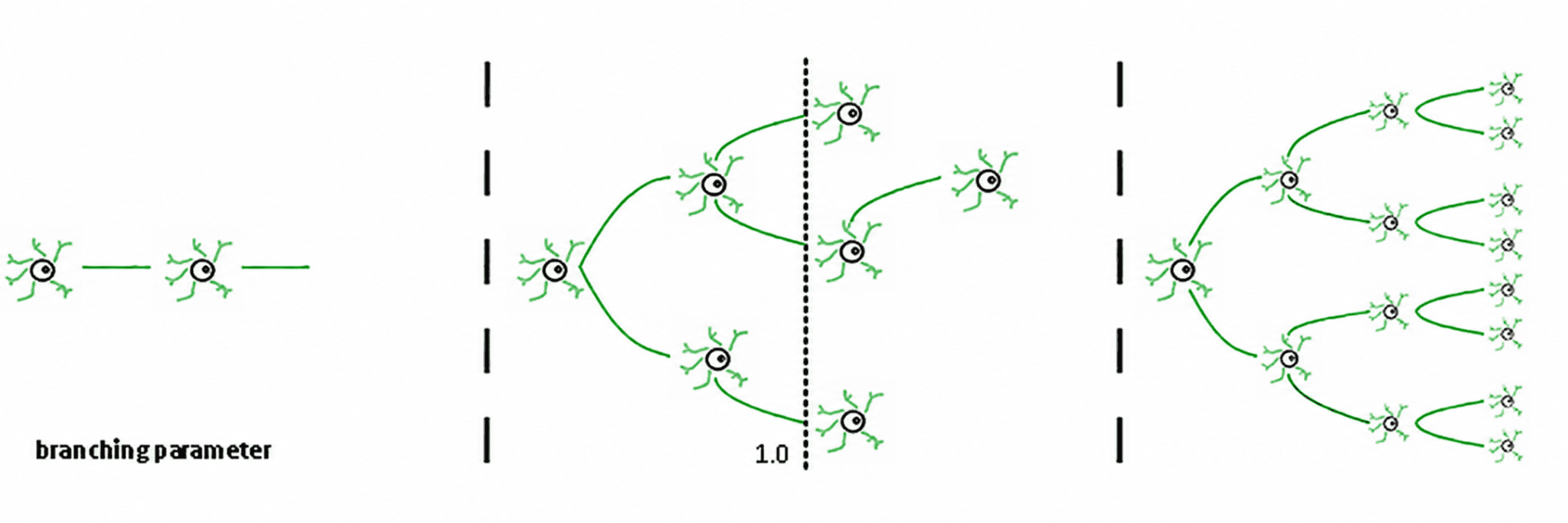
Trzy reżimy wynikają z tego (de Carvalho & Prado, 2000; Haldeman & Beggs, 2005):
Podkrytyczny (σ < 1): aktywność zanika; system szybko "zapomina" perturbację. Jest stabilny, ale słaby w pamięci i mało ekspresyjny.
Nadkrytyczny (σ > 1): aktywność eksploduje w kaskady. To jest sygnatura patologicznych reżimów, takich jak napady epileptyczne (Hsu i in., 2008; Hagemann i in., 2021).
Krytyczny (σ ≈ 1): każdy wystrzał, w średniej, generuje kolejny wystrzał. Aktywność reverberuje, lawiny neuronalne obey power laws, a system utrzymuje uporządkowaną pamięć wejściową.
Piękno σ polega na tym, że jest to pojedyncza liczba, która podsumowuje globalny reżim dynamiczny. Ale jego pomiar jest mniej trywialny. Gdy zastosowane do nagrań korowych in vivo, pomiar ujawnia, że kora nie działa dokładnie przy σ = 1, ale nieco poniżej, w reżimie, który autorzy nazywają reverberującym (Wilting i in., 2018). Różnica jest ważna: bycie dokładnie przy σ = 1 byłoby jak pedałowanie roweru balansującego na linie; bycie nieco poniżej pozwala na szybką regulację w odpowiedzi na wymagania zadania bez ryzyka eksplozji.
Krytyczność w sztucznych sieciach neuronowych: Od krawędzi chaosu do obliczeń w zbiorniku
Bertschinger i Natschläger (2004) wykazali, że losowe recurrent threshold networks osiągają swoją maksymalną zdolność obliczeniową w zadaniach przetwarzania temporalnego dokładnie w momencie przejścia od porządku do chaosu.
Boedecker i in. (2012) rozszerzyli analizę na sieci echo state w ramach paradygmatu obliczeń w zbiorniku, potwierdzając, że pojemność transferu informacji i aktywna pamięć są maksymalizowane na krawędzi chaosu.
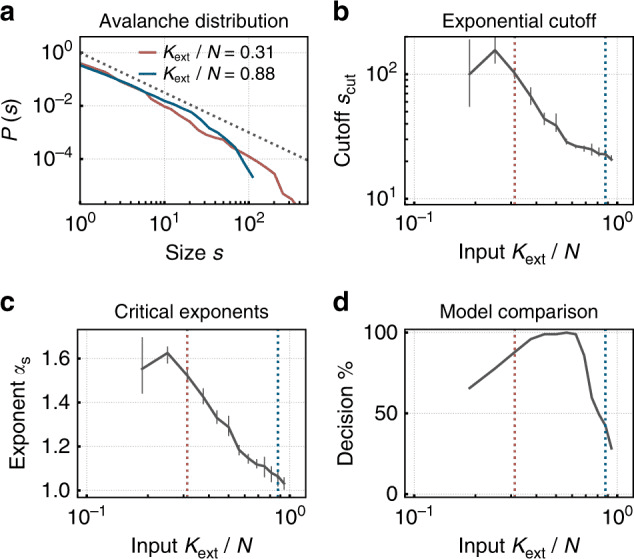
Rys. 3. Sieć neuromorficzna ze skokową plastycznością samoorganizuje się w kierunku krytyczności przy niskim zewnętrznym wejściu, pokazując rozkłady rozmiarów lawin na zasadzie potęgowej — znak stanu krytycznego zarówno w biologicznych, jak i sztucznych sieciach neuronowych. Przy wyższym wejściu, sieć przesuwa się w reżim podkrytyczny z przyciętymi rozkładami. Odtworzone z Cramer i in. (2020), Nature Communications, 11, 2853. CC BY 4.0.
W języku sztucznych sieci neuronowych, parametr pomiarowy nazywany jest promieniem spektralnym. Gdy przekracza 1, trajektorie rozdzielają się wykładniczo (chaos); gdy jest znacznie poniżej 1, sieć zapada się do punktu stałego i traci pamięć. Promień spektralny bliski 1 jest w tym kontekście formalnym odpowiednikiem biologicznego σ ≈ 1 (Magnasco, 2022; Morales i in., 2023). W sieciach neuronowych o skokowych, wskaźnik rozgałęzienia można mierzyć metodami prawie identycznymi do tych używanych w kulturach neuronowych (Cramer i in., 2020; Zeraati i in., 2024).
Dlaczego krytyczność mózgu maksymalizuje obliczenia neuronowe?
Działanie w pobliżu σ ≈ 1 zapewnia cztery korzyści, które są centralne zarówno dla hipotezy krytyczności mózgu, jak i dla projektowania systemów AI inspirowanych mózgiem:
Maksymalny zakres dynamiczny. Shew i in. (2009) wykazali, że zakres intensywności wejściowych, które kora może rozróżnić, jest maksymalny, gdy równowaga ekscytacji i hamowania umieszcza sieć w stanie krytycznym.
Maksymalna pojemność informacyjna. Entropia wzorców lawinowych i wspólna informacja między wejściem a wyjściem osiągają szczyt przy σ ≈ 1 (Shew i in., 2011).
Optymalne pamięć zanikająca. W reżimie krytycznym, perturbacja jest podtrzymywana wystarczająco długo, aby wpłynąć na przetwarzanie bez zanieczyszczania odległej przyszłości; to idealne miejsce pomiędzy stabilnością a integracją czasową (Boedecker i in., 2012).
Złożoność jako miara jednocząca. Timme i in. (2016) wykazali, że złożoność neuronowa jest maksymalizowana dokładnie w punkcie krytycznym, łącząc krytyczność z formalnymi teoriami świadomości i przetwarzania.
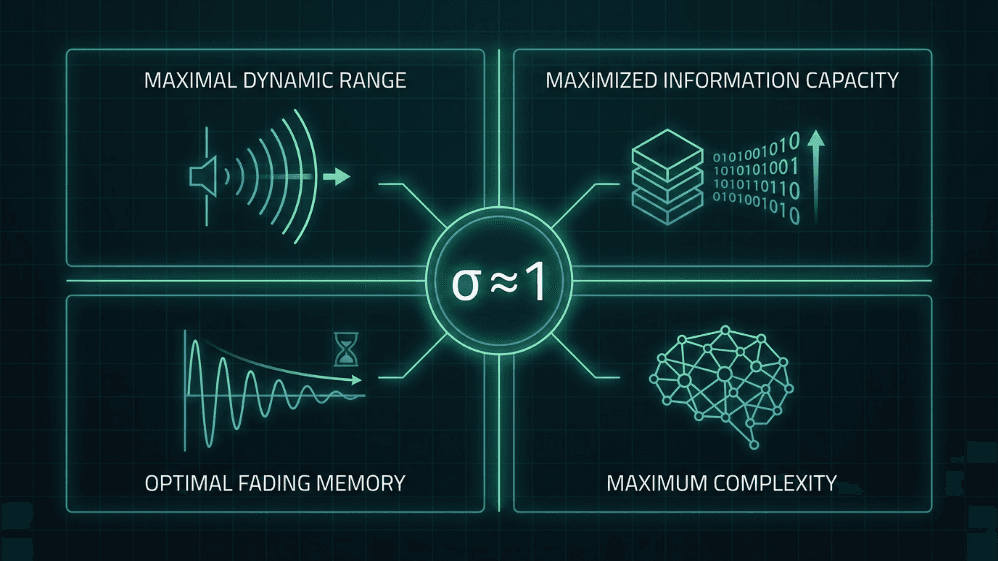
Rys. 4. Cztery zalety obliczeniowe działania w pobliżu krytycznego wskaźnika rozgałęzienia (σ ≈ 1). W stanie krytycznym sieci neuronowe osiągają maksymalny zakres dynamiczny, maksymalną pojemność informacyjną, optymalną pamięć zanikającą oraz maksymalną złożoność — właściwości, które są centralne zarówno dla hipotezy krytyczności mózgu, jak i dla projektowania AI inspirowanej mózgiem.
Mózg nie zawsze działa przy σ = 1
To nie oznacza, że mózg zawsze działa przy σ = 1. Dowody raczej sugerują nieco podkrytyczny i modulowany reżim: podczas wymagających zadań sieć zbliża się do krytyczności, podczas głębokiego snu oddala się, a stany patologiczne (epilepsja, głęboka anestezja, niektóre stany psychiatryczne) są związane z mierzalnymi odchyleniami od tego zakresu operacyjnego (Meisel i in., 2017; Zimmern, 2020). Wskaźnik rozgałęzienia staje się dynamicznym biomarkerem funkcjonalnego stanu układu nerwowego.
Dlaczego używamy wskaźnika rozgałęzienia w Neuraxon: projekt AI inspirowany biologią na krawędzi chaosu
Neuraxon jest systemem inspirowanym biologią, który przyjmuje dynamiczne zasady kory jako ograniczenia projektowe. Wskaźnik rozgałęzienia jest jednym z najważniejszych, a używamy go z czterech powodów:
Jako operacyjny inwariant w czasie rzeczywistym dla stabilności sieci neuronowych
W głębokich architekturach skokowych lub recurrent, podwójne ryzyko załamania aktywności (cicha sieć, zanikające gradienty) oraz eksplozji (nasycenie, eksplodujące gradienty) jest strukturalne. Monitorowanie σ w czasie rzeczywistym daje nam pojedynczy diagnostyczny skalar, niezależny od konkretnej architektury, który wskazuje, czy system żyje w sensie obliczeniowym.
Jako cel samoregulacji inspirowany biologią przez samoorganizującą się krytyczność
Sieć samoorganizuje się w kierunku krytyczności bez potrzeby centralnego dostrajania, replikując zasadę samoorganizującej się krytyczności (Bornholdt & Röhl, 2003; Levina i in., 2007). To drastycznie redukuje wrażliwość na hiperparametry i nadaje systemowi odporność na zmiany rozkładu. Jak zbadaliśmy w Tomie 7 NIA na temat życia sztucznego i ekosystemów cyfrowych, dokładnie w ten sposób powstaje złożoność emergentna z lokalnych reguł bez centralnej kontroli.
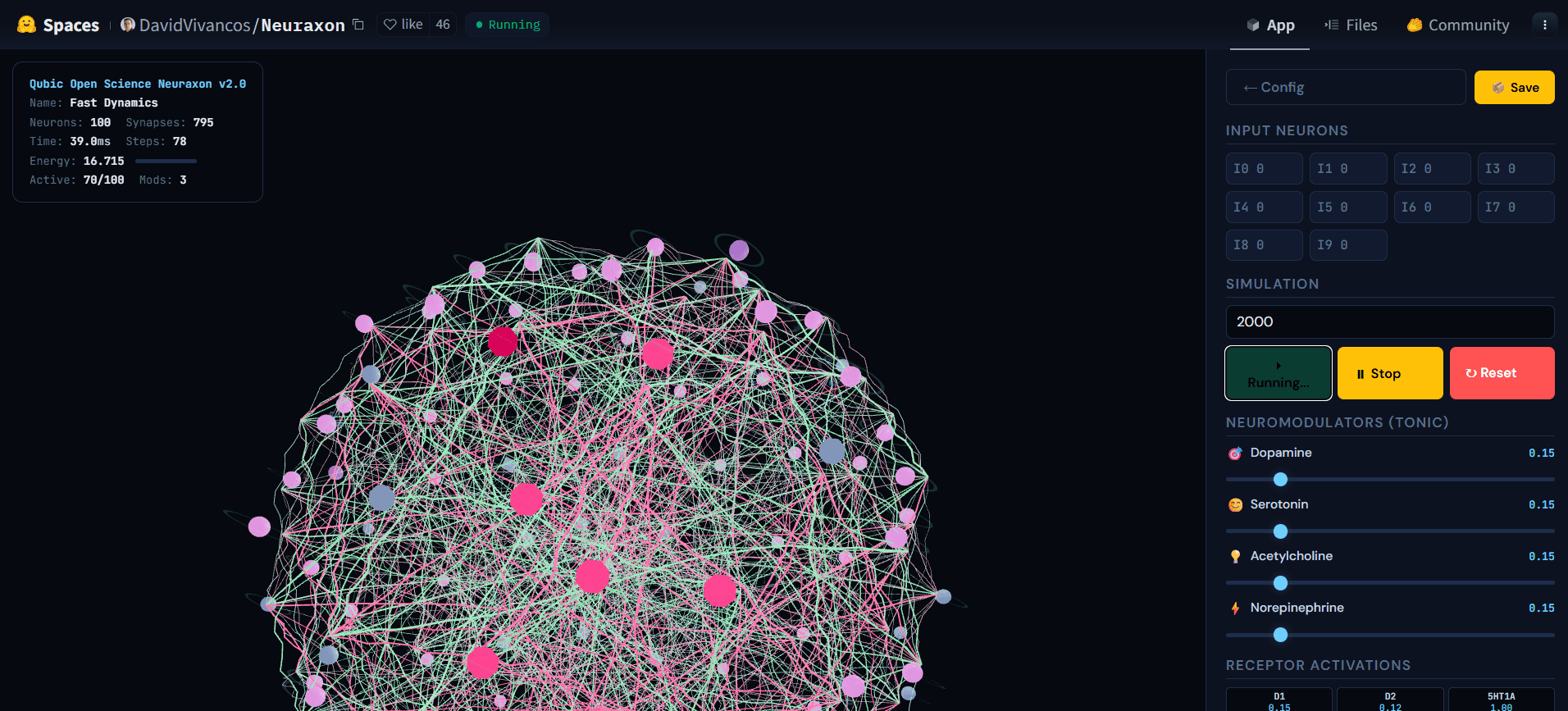
Rys. 5. Sieć 3D Neuraxon podczas aktywnej symulacji, pokazująca kaskadową aktywność w neuronach o ternarnym stanie. Jasno aktywne węzły (różowy) propagują sygnały przez połączenia ekscytujące (zielony) i hamujące (różowy), podczas gdy inne neurony pozostają w spoczynku (szary), ilustrując reżim reverberacyjny w pobliżu krytycznego wskaźnika rozgałęzienia (σ ≈ 1). Ten zrównoważony stan — ani cichy, ani eksplodujący — jest tym, do czego Neuraxon samoorganizuje się, stosując zasady krytyczności inspirowane biologią. Zbadaj interaktywną demonstrację na huggingface.co/spaces/DavidVivancos/Neuraxon. Źródło: Zespół Naukowy Qubic.
Jako most między obserwacją neurobiologiczną a projektowaniem AI
Wskaźnik rozgałęzienia jest jedną z bardzo nielicznych wielkości, które są mierzone tym samym formalizmem w elektrofizjologii, fMRI i sieciach sztucznych. Umożliwia to testowanie hipotez dwukierunkowych: jeśli interwencja poprawia biologiczną krytyczność, możemy zapytać, czy ta sama interwencja — przetłumaczona na architekturę sztuczną — poprawia obliczenia modelu, i odwrotnie. Ta zasada jest centralna dla ram neuromodulacji oraz mechanizmów bramkowania astrocytów, które opracowaliśmy w poprzednich tomach tej akademii.
Jako kryterium funkcjonalne, a nie estetyczne, dla AI inspirowanej mózgiem
Krytyczność jest operacyjnym ograniczeniem z empirycznymi konsekwencjami. Działanie w pobliżu reżimu reverberacyjnego poprawia — jak mierzone w naszych wewnętrznych ocenach i zgłoszonych publikacjach — zdolność do generalizacji, stabilność w obliczu perturbacji wejściowych, bogactwo reprezentacyjną oraz spójność czasową rozumowania. Te efekty jakościowo odpowiadają tym zgłoszonym zarówno w literaturze biologicznej (Cocchi i in., 2017), jak i sztucznej (Cramer i in., 2020; Morales i in., 2023).
Wskaźnik rozgałęzienia: od fizyki statystycznej do architektury AI inspirowanej mózgiem
Wskaźnik rozgałęzienia jest jednym z tych koncepcyjnych rara avis: wystarczająco prosty, by sprowadzić się do pojedynczej formuły, wystarczająco głęboki, by łączyć fizykę statystyczną, neuronaukę, AI i projektowanie systemów. Dla mózgu biologicznego, σ ≈ 1 wydaje się być reżimem, w którym pojawia się cnotliwa kombinacja wrażliwości, pamięci, ekspresyjności i odporności. Dla sztucznych sieci ta sama granica — przemianowana na krawędź chaosu — przewiduje maksymalną zdolność obliczeniową.
A dla Neuraxon, to jest zasada przewodnia projektowania inspirowanego biologią: audytowalny, samoregulujący się i biologicznie znaczący wskaźnik, który pomaga nam utrzymać system przy życiu, w najszerszym znaczeniu tego słowa.
Bibliografia
Beggs, J. M., & Plenz, D. (2003). Lawiny neuronalne w obwodach neokortykalnych. The Journal of Neuroscience, 23(35), 11167–11177. https://doi.org/10.1523/JNEUROSCI.23-35-11167.2003
Bertschinger, N., & Natschläger, T. (2004). Obliczenia w czasie rzeczywistym na krawędzi chaosu w recurrent neural networks. Neural Computation, 16(7), 1413–1436. https://doi.org/10.1162/089976604323057443
Boedecker, J., Obst, O., Lizier, J. T., Mayer, N. M., & Asada, M. (2012). Przetwarzanie informacji w sieciach echo state na krawędzi chaosu. Theory in Biosciences, 131(3), 205–213. https://doi.org/10.1007/s12064-011-0146-8
Bornholdt, S., & Röhl, T. (2003). Samoorganizujące się krytyczne sieci neuronowe. Physical Review E, 67(6), 066118. https://doi.org/10.1103/PhysRevE.67.066118
Cocchi, L., Gollo, L. L., Zalesky, A., & Breakspear, M. (2017). Krytyczność w mózgu: synteza neurobiologii, modeli i poznania. Postępy w Neurobiologii, 158, 132–152. https://doi.org/10.1016/j.pneurobio.2017.07.002
Cramer, B., Stöckel, D., Kreft, M., Wibral, M., Schemmel, J., Meier, K., & Priesemann, V. (2020). Kontrola krytyczności i obliczenia w skokowych sieciach neuromorficznych z plastycznością. Nature Communications, 11, 2853. https://doi.org/10.1038/s41467-020-16548-3
de Carvalho, J. X., & Prado, C. P. C. (2000). Samoorganizująca się krytyczność w modelu Olami-Feder-Christensen. Physical Review Letters, 84(17), 4006–4009. https://doi.org/10.1103/PhysRevLett.84.4006
Derrida, B., & Pomeau, Y. (1986). Losowe sieci automatów: prosta przybliżona wersja. Europhysics Letters, 1(2), 45–49. https://doi.org/10.1209/0295-5075/1/2/001
Hagemann, A., Wilting, J., Samimizad, B., Mormann, F., & Priesemann, V. (2021). Ocena krytyczności w aktywności pojedynczych neuronów przed napadami w ludzkiej korze epileptycznej. PLOS Computational Biology, 17(3), e1008773. https://doi.org/10.1371/journal.pcbi.1008773
Haldeman, C., & Beggs, J. M. (2005). Krytyczne rozgałęzianie uchwyca aktywność w żywych sieciach neuronowych i maksymalizuje liczbę metastabilnych stanów. Physical Review Letters, 94(5), 058101. https://doi.org/10.1103/PhysRevLett.94.058101
Hsu, D., Chen, W., Hsu, M., & Beggs, J. M. (2008). Otwarte hipoteza: czy epilepsja jest nauczana i czy można ją oduczyć? Epilepsja & Zachowanie, 13(3), 511–522. https://doi.org/10.1016/j.yebeh.2008.05.007
Langton, C. G. (1990). Obliczenia na krawędzi chaosu: przejścia fazowe i emergentne obliczenia. Physica D: Nonlinear Phenomena, 42(1–3), 12–37. https://doi.org/10.1016/0167-2789(90)90064-V
Levina, A., Herrmann, J. M., & Geisel, T. (2007). Dynamiczne synapsy powodujące samoorganizującą się krytyczność w sieciach neuronowych. Nature Physics, 3(12), 857–860. https://doi.org/10.1038/nphys758
Magnasco, M. O. (2022). Odporność i elastyczność funkcji neuronowych dzięki dynamice krytycznej. Entropia, 24(5), 591. https://doi.org/10.3390/e24050591
Meisel, C., Klaus, A., Vyazovskiy, V. V., & Plenz, D. (2017). Interakcja pomiędzy długozasięgowymi i krótkozasięgowymi korelacjami czasowymi kształtuje dynamikę kory mózgowej w różnych stanach czuwania. The Journal of Neuroscience, 37(42), 10114–10124. https://doi.org/10.1523/JNEUROSCI.0448-17.2017
Morales, G. B., di Santo, S., & Muñoz, M. A. (2023). Odkrywanie wewnętrznej dynamiki biologicznych i sztucznych sieci neuronowych: Od krytyczności do optymalnych reprezentacji. Frontiers in Complex Systems, 1, 1276338. https://doi.org/10.3389/fcpxs.2023.1276338
Plenz, D., Ribeiro, T. L., Miller, S. R., Kells, P. A., Vakili, A., & Capek, E. L. (2021). Krytyczność samoorganizująca się w mózgu. Frontiers in Physics, 9, 639389. https://doi.org/10.3389/fphy.2021.639389
Shew, W. L., Yang, H., Petermann, T., Roy, R., & Plenz, D. (2009). Lawiny neuronalne sugerują maksymalny zakres dynamiczny w sieciach korowych w stanie krytycznym. The Journal of Neuroscience, 29(49), 15595–15600. https://doi.org/10.1523/JNEUROSCI.3864-09.2009
Shew, W. L., Yang, H., Yu, S., Roy, R., & Plenz, D. (2011). Pojemność informacyjna i transmisja są maksymalizowane w zrównoważonych sieciach korowych z lawinami neuronalnymi. The Journal of Neuroscience, 31(1), 55–63. https://doi.org/10.1523/JNEUROSCI.4637-10.2011
Spitzner, F. P., Dehning, J., Wilting, J., Hagemann, A., Neto, J. P., Zierenberg, J., & Priesemann, V. (2021). MR. Estimator, narzędzie do określania wewnętrznych czasów na podstawie podpróbkowanej aktywności skokowej. PLOS ONE, 16(4), e0249447. https://doi.org/10.1371/journal.pone.0249447
Timme, N. M., Marshall, N. J., Bennett, N., Ripp, M., Lautzenhiser, E., & Beggs, J. M. (2016). Krytyczność maksymalizuje złożoność w tkankach neuronowych. Frontiers in Physiology, 7, 425. https://doi.org/10.3389/fphys.2016.00425
Turrigiano, G. G. (2008). Samostrojący się neuron: skalowanie synaptyczne synaps ekscytujących. Cell, 135(3), 422–435. https://doi.org/10.1016/j.cell.2008.10.008
Wang, J., Cao, R., Brunton, B. W., Smith, R. E. W., Buckner, R. L., & Liu, T. T. (2025). Genetyczne wkłady w krytyczność mózgu i jej związek z ludzkimi funkcjami poznawczymi. Proceedings of the National Academy of Sciences, 122(26), e2417010122. https://doi.org/10.1073/pnas.2417010122
Wilting, J., Dehning, J., Pinheiro Neto, J., Rudelt, L., Wibral, M., Zierenberg, J., & Priesemann, V. (2018). Działanie w reżimie reverberacyjnym umożliwia szybką regulację stanów sieci w odpowiedzi na wymagania zadania. Frontiers in Systems Neuroscience, 12, 55. https://doi.org/10.3389/fnsys.2018.00055
Wilting, J., & Priesemann, V. (2018). Wnioskowanie zbiorowych stanów dynamicznych z szeroko nieobserwowanych systemów. Nature Communications, 9, 2325. https://doi.org/10.1038/s41467-018-04725-4
Wilting, J., & Priesemann, V. (2019). 25 lat krytyczności w neuronauce — Ustanowione wyniki, otwarte kontrowersje, nowe koncepcje. Current Opinion in Neurobiology, 58, 105–111. https://doi.org/10.1016/j.conb.2019.08.002
Yu, C. (2022). W kierunku zjednoczonej analizy hipotezy krytyczności mózgu: przegląd kilku dostępnych narzędzi. Frontiers in Neural Circuits, 16, 911245. https://doi.org/10.3389/fncir.2022.911245
Zeraati, R., Engel, T. A., & Levina, A. (2024). Szacowanie wewnętrznych czasów i krytyczności z nagrań neuronowych: Metody i pułapki. Current Opinion in Neurobiology, 86, 102871. https://doi.org/10.1016/j.conb.2024.102871
Zimmern, V. (2020). Dlaczego krytyczność mózgu jest klinicznie istotna: przegląd. Frontiers in Neural Circuits, 14, 54. https://doi.org/10.3389/fncir.2020.00054
Zbadaj pełną Akademię Inteligencji Neuraxon
To jest Tom 8 z #Neuraxon Inteligencji #academy zespołu #Qubic naukowego. Jeśli dopiero do nas dołączasz, zbadaj pełną serię, aby zbudować pełne zrozumienie nauki stojącej za Neuraxon, #aigarth i podejściem Qubic do inspirowanej mózgiem, #decentralized sztucznej inteligencji:
NIA Vol. 1: Dlaczego inteligencja nie jest obliczana w krokach, ale w czasie — Wyjaśnia, dlaczego inteligencja biologiczna działa w czasie ciągłym, a nie w dyskretnych krokach obliczeniowych jak tradycyjne LLM.
NIA Vol. 2: Dynamika ternarna jako model inteligencji żywej — Wyjaśnia dynamikę ternarną i dlaczego logika trójskalowa (ekscytująca, neutralna, hamująca) ma znaczenie w modelowaniu systemów żywych.
NIA Vol. 3: Neuromodulacja i AI inspirowana mózgiem — Obejmuje neuromodulację i jak chemiczne sygnalizowanie mózgu (dopamina, serotonina, acetylocholina, norepinefryna) inspiruje architekturę Neuraxona.
NIA Vol. 4: Sieci neuronowe w AI i neuronauce — Głębokie porównanie biologicznych sieci neuronowych, sztucznych sieci neuronowych oraz trzeciej ścieżki Neuraxona.
NIA Vol. 5: Astrocyty i AI inspirowana mózgiem — Jak bramkowanie astrocytów przekształca plastyczność sieci neuronowych poprzez ramy AGMP w Neuraxon.
NIA Vol. 6: Świadome maszyny vs inteligentne organizmy: Wyjaśniona świadomość AI — Bada świadomość AI przez pryzmat Teorii Global Workspace, Teorii Zintegrowanej Informacji oraz kodowania predykcyjnego.
NIA Vol. 7: Gra życia Conwaya, życie sztuczne i ekosystemy cyfrowe — Nauka stojąca za Qubic, Aigarth i podejściem Neuraxona do emergentnej złożoności i samoorganizującej się krytyczności w zdecentralizowanej AI.
Qubic to zdecentralizowana, otwarta sieć dla technologii eksperymentalnej. Aby dowiedzieć się więcej, odwiedź qubic.org. Dołącz do dyskusji na X, Discord i Telegram.